5h du matin Lucile, en stage à l’ONF, vient me chercher et on part en direction du monastère de la Grande Chartreuse. On y rejoint Sylvain, agent du réseau Avifaune de l’ONF, qui procède depuis quelques semaines à des inventaires ornithologiques dans différentes régions françaises.
Après un peu de grimpe dans les forêts peu entretenues au dessus du monastère, nous arrivons à une placette (point GPS choisi préalablement et au hasard). Nous y restons 20 minutes et Sylvain note les différentes espèces d’oiseaux qu’il identifie au chant et aux jumelles. On recommence ce protocole à différents endroits. C’est la répétition des passages des naturalistes sur ces points d’écoute qui permettra de confirmer les observations. Les informations récoltées sont reportées dans la base de données naturalistes de l’ONF et transmises à l’INPN
7h On se dirige vers un spot où des rougequeues à front blanc (Phoenicurus phoenicurus) auraient été observés. Cette espèce est présente en France entre fin mars et début septembre, moment où il repart dans ses quartiers d’hivernage, en Afrique sahélienne. Bingo ! On entend son chant puis on arrive à l’observer en bordure de champs dans un petit village.
8h Un café avalé, Lucile doit repartir et les agents de l’ONF me proposent de les suivre sur les crêtes au dessus de St Pierre de Chartreuse, à la recherche de rapaces. Proposition que je ne peux pas refuser. Et me voilà, cavalant à l’improviste les crêtes du massif de la Chartreuse, sur les traces des rapaces (et à la recherche de crottes de gélinotte) et à la rencontre des chamois pendant le reste de la journée.
Merci à Lucile, Thierry et Sylvain pour cette journée.
Le protocole IPA. La méthode des Indices Ponctuels d’Abondance a été élaborée et décrite par Blondel, Ferry et Frochot en 1970 . Cette méthode consiste, aux cours de deux sessions distinctes de comptage, à noter l’ensemble des oiseaux observés ou entendus durant 20 minutes à partir d’un point fixe du territoire.
4h du matin. Réveil silencieux au milieu des équipes de bénévoles qui vivent ensemble dans un lieu gardé secret. Il est temps d’enfiler plusieurs couches de vêtements et d’embarquer les combinaisons étanches dans la voiture. Une heure et quelques dizaines de kilomètres plus tard, nous arrivons au port pour prendre la relève de ceux qui sont restés toute la nuit afin de veiller sur le bateau. En effet, au port il faut se faire discret, Sea Shepherd semble être considéré comme l’ennemi numéro 1 des pêcheurs. Cette sensation est confirmée par les différents messages radios interceptés et qui nous sont directement adressés durant la matinée. Au milieu de quelques messages cordiaux, bien que tendus, on nous interpelle, nous dit de dégager.
Des premières lueurs du jour jusqu’au milieu de l’après-midi, à 5 sur le zodiac, nous suivront les bateaux de pêche qui relèvent leurs filets. La mission est simple : rester à une certaine de distance de sécurité et filmer les remontées de filet. Les images récupérées permettent de comptabiliser et de témoigner du nombre de « prises accidentelles » de dauphins dans ces filets. Selon Pelagis, un observatoire du CNRS, de janvier à mars 2020, le RNE (Réseau National Échouages) a enregistré 1067 échouages de petits cétacés sur la façade atlantique et en Manche. D’après les examens réalisés, la capture dans un engin de pêche reste la principale cause de mortalité observée chez le dauphin commun lors des événements d’échouages multiples en hiver et ce depuis les années 1990.
Je me surprends à regarder l’horizon, vide, dans le bruit sourd des enrouleurs de lignes, les mouettes s’affairent autour des poissons que les pêcheurs rejettent à l’eau. Parfois le bruit d’un marteau qui s’obstine à détruire un crabe ou un coquillage emmêlé dans les fils me terrifie. Je pensais avoir conscience de la réalité de la pêche, j’ai grandi sur la côte atlantique et il m’est arrivé de pêcher à la ligne avec des copains ou de la famille. Mais je n’avais jusqu’à ce jour pas conscience de la violence. Entre les humains et les autres animaux, leurs besoins respectifs de (sur)vivre. La violence d’un système, d’un monde, d’une lutte.
Mon cœur se serre quand je me rends compte que nous suivons ce bateau depuis plusieurs heures et qu’il ne cesse de remonter ses entrailles de plastiques. Et sur la carte GPS, j’en vois des centaines. Reste-il une seule langue de fond marin qui ne soit recouverte de filet ?
Il y a un univers entier qui sépare les deux bords – à tribord, le soleil qui épouse doucement les vagues argentées, la chaleur qui vient sécher l’écume sur mon visage et mon appareil photo, le goût salé de l’infini qui s’offre à moi – à bâbord, les deux activistes qui luttent pour tenir debout et filmer sans interruption la remonté des filets, éblouis par le projecteur que les pêcheurs ont dirigé vers nous, les craquements, le vacarme mais en même temps le silence, lourd, des militant.e.s qui savent ce qui se jouent sous leurs yeux. Un énième acte d’une tragédie qui prend place en Atlantique.
Des militant.e.s qui sont avant tout, des hommes et des femmes, qui partagent une envie : que la mer reste vivante.
Je suis ravie d’annoncer que « Bestioles » sera exposée au festival OFF de Visa pour l’image à partir du 28 août 2021.
Bestioles
Ils rampent, ils grouillent, ils se cachent, ils nous effraient. Araignées, serpents, tritons ou insectes, ces petites bêtes sont essentiels au bon fonctionnement des écosystèmes. Inféodées aux Pyrénées et au Sud de la France, les modifications apportées à leur environnement déciment leurs populations, déjà menacées. Au-delà de leur image repoussante, ces bestioles permettent entre autres de réguler les populations de ravageurs (insectes ou petits mammifères abimant les cultures). Indispensables à l’équilibre naturel, ce sont des trésors régionaux.
Dans le cadre d’un Diplôme Universitaire de Photojournalisme, Captation et Images aériennes à l’Université de Perpignan, nous sommes partis dans la campagne de Marmande à la rencontre de André, un agriculteur ambitieux et pionnier de la culture de fruits à coque et de poivres en France.
Ses voisins le prennent pour un fou, mais son assurance et son investissement pourraient en convaincre plus d’un. André a passé sa vie à travailler dans les champs de maïs du Lot-et-Garonne. À la suite d’un accident lié au produit utilisé en agriculture conventionnelle, André et ses fils ont pris un virage à 90 degrés pour se lancer dans l’agriculture biologique. Le réchauffement climatique impacte fortement la vie des agriculteurs, André, veut y faire face et s’adapter en optant pour une production de fruits à coques, moins consommateurs d’eau que d’autres plantations. Pour s’y faire, il compte produire ces plantations lui-même, de l’engrais à la vente.
Baleine sous Gravillon (BSG) est un né le 21 mai 2020. Ce podcast natif et indépendant, de vulgarisation scientifique, ludique, non lucratif et bénévole, sans pub et grand public est entièrement consacré à des êtres vivants étonnants, connus ou méconnus, à la biodiversité et à la nature. J’ai rejoint Marc, la voix de BSG, dans cette aventure le 12 octobre 2020 et j’ai opéré en silence sous le totem de Cistude.
La saison 1 était consacrée au biomimétisme. Le podcast couvre désormais tous les sujets liés à la nature et à l’environnement. Dans la saison 2, qui a commencé le 2 septembre 2020, on retrouve des entretiens avec des personnalités, auteurs, scientifiques, réalisateurs, voyageurs, explorateurs, guides et photographes animaliers, sportifs, environnementalistes, artistes, dirigeants d’ONG ou d’entreprises… des « passeurs/passeuses » de savoirs et de messages.
Désormais BSG est une association qui propose sur son site internet les épisodes de BSG et des 3 petits frères de BSG (3 nouveaux podcasts : Combats, PPDP, Nomen), différentes publications (articles de vulgarisation, anecdotes etc.) et une galerie de photographes partenaires.
Étude des populations de serpents en Réserve Biologique Intégrale
CEBC/CNRS, Deux-Sèvres, France
Le Centre d’Etudes Biologiques de Chizé est une station de terrain du CNRS au cœur de la forêt de Chizé. La Réserve Biologique domaniale Intégrale (RBI) de la Sylve d’Argenson, en forêt de Chizé, a été créée en 2006. Dans une RBI, l’exploitation forestière est proscrite, ce qui permet aux scientifiques d’en étudier l’évolution naturelle.
L’équipe ECOPHY du CNRS étudie comment les animaux répondent aux stresseurs environnementaux (changement climatique, dégradation des habitats, pollution). L’une de leurs missions est d’effectuer un protocole de Capture-Marquage-Recapture (CMR) permettant d’estimer le nombre d’individus d’une population et d’en suivre l’évolution. Ce protocole est tout à fait adapté aux serpents : il ne modifie ni leur biologie, ni leur succès reproducteur ou leur taux de mortalité et il n’influe pas sur leurs déplacements.
Les espèces concernées par cette étude sont la couleuvre d’esculape (Zamesis longissimus), la couleuvre verte et jaune (Hierophis viridiflavus), la couleuvre helvétique (Natrix helvetica) et la vipère aspic (Vipera aspis).
La capture
Les serpents sont des animaux diurnes mais très discrets. Leur observation est donc difficile sans méthode de capture.
L’équipe a donc installé des plaques en fibrociment ondulées de 1m², sous lesquelles les serpents viennent se cacher pour se thermoréguler en sécurité. Ces plaques ne sont soulevées qu’une fois par semaine, afin que l’étude ne soit pas trop invasive.
Chaque jour un des différents parcours est effectué par Martin, qui note précisément l’emplacement , l’heure, le numéro de la plaque sous laquelle l’animal a été trouvé ainsi que son espèce. Il se déplace muni de tout son matériel de capture (crochets, gants de sécurité, sacs de transport).
L’identification de l’individu doit se faire rapidement avant sa capture, afin de prendre les dispositions nécessaires. En effet, s’il s’agit d’une vipère aspic, une espèce venimeuse, la méthode de capture est différente et des précautions doivent être prises.
Pour attraper une vipère sans la blesser et limiter son stress, on utilise un crochet la mettre directement dans la boîte à clapet, boite sécurisée et fermés. On indique sur cette boite la présence d’un animal venimeux afin de ne pas la confondre avec les autres boites, où sont déposés les sacs en tissus contenant les couleuvres.
Les animaux sont amenés au laboratoire, où ils passeront la nuit au chaud dans les boîtes conçues pour empêcher les évasions.
Martin attrapant une couleuvre d’Esculape (Zamenis longissimus) sous l’une des plaques numérotées de la Réserve. En été, les captures se font en fin de journée, quand la température est favorable (environ 20°C).
Les serpents capturés sont transportés dans des sacs en tissus opaques et placés dans une boite à clapet fermée.
Les mesures biométriques
Une fois au laboratoire, une fiche biométrique doit être remplie pour chaque individu.
Si l’individu est déjà marqué, on peut l’identifier et retrouver sa fiche, avec les données mesurées lors de sa dernière capture. S’il n’a jamais été capturé, l’individu doit être marqué. On marque les écailles correspondantes au numéro de l’individu dans le répertoire au fer à souder. Cette technique de marquage possède un taux d’erreur d’identification (2%) plus faible que la puce (5%) qui peut être perdue dans le corps de l’animal si celle-ci est mal implantée.
Mesure de la taille de la mâchoire (Zamenis longissimus) avec un pied à coulisse.Mesure de la taille du serpent (Zamenis longissimus)
Chaque fiche biométrique est accompagnée d’une photo(copie) en vue ventrale, afin d’aider à l’identification.Mesure de la largeur du crâne (Zamenis longissimus)
Prise des mesures sur une couleuvre d’Esculape (Zamenis longissimus).
Un numéro est attribué à chaque individu et un léger marquage au fer à souder sur les écailles est réalisé. Ce marquage est indolore et plus fiable que l’identification par puce.
Pour avoir des renseignements sur le régime alimentaire de chaque espèce, on récolte les fèces que l’on dilue dans l’alcool pour voir le contenu du dernier repas (poils, plumes, écailles, élytres).
Vipère aspic (Vipera aspis) juvénile, première capture pour ce jeune spécimen de l’année dernière.Vipère aspic (Vipera aspis) juvénile,. Elle est placée dans un terrarium avec eau et chaleur.
Les relâchers
Une fois les données récoltées, les serpents capturés sont relâchés sous la plaque où ils ont été trouvés.
Étudier les animaux et effectuer un suivi sur le long terme (une dizaine d’année pour cette CMR) est un travail à plein temps. Les prospections et les captures/relâchés peuvent durer jusqu’à tard, notamment quand le chemin est difficile. Les réserves biologiques intégrales ne sont pas gérées par des forestiers, et le parcours est parfois peu dégagé, les plaques difficiles à trouver pour les regards non-avertis. Ce sont en général des stagiaires, comme Martin, issus de BTS ou de cursus universitaires, qui récoltent et analysent le premier jet des données.
Les petites mains de la science ne signent pas les papiers qui paraissent dans les revues, ils ne sont souvent pas rémunérés et sont là par pure passion.
Couleuvre d’Esculape (Zamenis longissimus)
La nuit tombée, Martin finit son parcours à la frontale. À ces heures tardives, il est courant d’observer et d’entendre la faune nocturne qui s’active .
Les gants et le crochet permettent de soulever les plaques sous lesquelles peuvent se trouver des vipères.
Crapaud accoucheur (Alytes obstetricans)Les jolies rencontres de fin de soirée… Un grand merci à Martin et à l’équipe herpéto de terrain.
Travail de recherche réalisé dans le cadre d’un cours de Philosophie de la Biologie (Licence de Philosophie – Sorbonne Université)
Biologie de l’évolution et écologie, les concepts phares
Définitions et concepts en écologie
Le monde naturel, ou biosphère, est régi par un ensemble de liens tissés entre les êtres vivants, les entités virales et les facteurs abiotiques (chimiques, géologiques etc). Tout objet vivant ou non-vivant que l’on retrouve dans cette réalité naturelle est associé à d’autres objets et trouve ainsi sa place dans une sorte de « grand rouage » de la nature. Ce sont ces relations entre différentes entités biotiques ou abiotiques qui forment un ensemble d’écosystèmes, interdépendants, avec leur fonctionnement et leurs populations propres. L’écologie est donc la science qui s’occupe de l’étude de ces milieux où vivent les êtres vivants, ainsi que des rapports de ces êtres avec leur milieu.
Tous les organismes vivants dépendent de la conversion première de l’énergie en matière organique par les végétaux. Ceux-ci sont donc à la base des chaînes alimentaires mais ils dépendent de facteurs abiotiques comme la concentration en minéraux des sols, la luminosité, la teneur en dioxyde de carbone de l’atmosphère etc..
La diversité interne à la biosphère qui permet la formation de différents écosystèmes est en fait relative à la distribution spatiale et asymétriques des océans et des continents, de leurs reliefs hétérogènes; à la diversité dans la cotisation biogéochimique des sols et des eaux continentales, à la variabilité géographique et temporelle des climats, à l’hétérogénéité dans la répartition et la composition de la végétation, à l’abondance relative et à la diversité des espèces en fonction des milieux. Un espace caractérisé par des facteurs abiotiques communs (comme le climats, l’altitude, les concentrations en éléments biogènes) est appelé un biotope. Dans ce biotope coexiste un ensemble d’organismes nommé la biocénose. Chaque écosystème est donc formé par un biotope et une biocénose. Il n’y a pas de frontières strictes entre les différents écosystèmes, c’est pour cela qu’on parle d’organisation fonctionnelle des écosystèmes.
La répartition des espèces ne semble pas aléatoire, les organismes vivants constituent des systèmes complexes associés à des milieux déterminés dans lesquels on trouve des communautés spécifiques, des associations particulières d’organismes avec des distributions variables mais organisées. Les interactions entres ces organismes sont structurées, elles ont des effets positifs et négatifs plus ou moins directs et dépendent principalement du critère de survie. Le « but » de chaque composante de l’écosystème semblant être sa propre survie aux dépens des autres composantes, et ainsi, les interactions entres entités de l’écosystème sont principalement des interactions d’ordre trophique ou sexuel.
On peut étudier des écosystèmes sous plusieurs échelles. Le plus petite est celle de l’individu, nous parlerons alors ici d’un seul organisme vivant (réel ou fictif). C’est de l’ordre du particulier, on s’intéresse à tel spécimen c’est-à-dire tel exemplaire d’une espèce. Un ensemble d’organismes de même espèce vivant dans un milieu donné à un moment donné forme une population. Lorsque plusieurs populations d’espèces différentes coexistent dans un même écosystème on parle alors de communauté. Une métapopulation est un réseau de populations séparées dans l’espace mais interconnectées par des flux d’individus ou de propagules.
Nous ne nous étendrons pas sur la difficulté de définir le concept d’espèce mais il est essentiel de comprendre l’enjeu de cette définition et donc pour cela son histoire.
Le fixisme de Linné (1735) est le reflet d’une conception biblique du monde. Sa définition de l’espèce évolue dans un cadre fixiste et affirme que les espèces sont immuables, issues d’une création divine et que par là on peut les définir par des critères morphologiques. Le concept d’espèce alors dit typologique.
Avec Buffon (1749), le monde scientifique prend conscience qu’il peut y avoir une multitude de formes vivantes. On définit alors l’espèce en affirmant que des individus qu’appartiendront s’ils peuvent se croiser et produire une descendance féconde. On ne prend plus seulement en compte l’unité d’une forme ou d’un type mais on instaure une dynamique avec la dimension généalogique de l’idée qu’une espèce est une lignée. C’est le concept biologique de l’espèce. Dans cette définition, Buffon tient compte de la perpétuation de la collection d’individus de même espèce au cours du temps.
L’anatomiste Georges Cuvier rejoint la définition biologique posée par Buffon mais remarque que concernant les espèces fossiles, seule un définition typologique est utilisable. La définition biologique de l’espèce se heurte à sa théorie du catastrophisme. Au contraire de ces derniers, Saint-Hilaire, qui a effectué de travaux sur la comparaisons des embryons et des foetus, s’inscrit dans le courant transformisme (affirmant que les espèces ont pu subir des changements au cours du temps).
Le transformisme est marqué par les théories de Lamarck qui pense que l’espèce est naturelle et que les classes, les ordres, les familles et les genres sont des divisions arbitraires. La définition qu’il donne est typologique essentiellement morphologique.
Darwin avec sa théorie de l’évolution et l’Origine des Espèces (1859) apporte l’idée d’une descendance avec des modifications quant aux parents, dont le mécanisme moteur est la sélection naturelle. C’est l’idée d’une divergence à partir d’un même ancêtre, il introduit ici l’idée de parenté entre les espèces. Cependant Darwin montre peu d’intérêt quant à une définition formelle de l’espèce.
Enfin, la définition désormais la plus communément admise est celle établie par Mayr en 1942. C’est une conception biologique de l’espèce qui la définit comme une population ou un ensemble de population dont les individus peuvent, au moins potentiellement, se reproduire entre eux et engendre une descendance viable et fécondes.
« Suivez la nourriture et vous comprendrez la structure des communautés »
Pour comprendre la structure des communautés et donc des écosystèmes, selon Charles Elton – éminent et reconnu écologue et zoologiste britannique – il faut suivre les chaines alimentaires et les interactions trophiques entre composantes de la communautés. En disant cela, il marque l’émergence des concepts de chaines alimentaires, niches écologiques et réseaux trophiques. Une relation trophique est une interaction d’ordre alimentaire entre deux organismes dont l’un est la ressource et l’autre le consommateur. La biomasse circule de la ressource vers le consommateur. La chaine alimentaire est un emboitement de relations trophiques, elle est définie comme une série d’organismes dans laquelle un organisme consomme celui qui le précède avant d’être mangé par celui qui le suit. Un réseau trophique est un un entrelacs de chaines alimentaires, un assemblage d’espèces d’un même écosystème réunies par les relations consommé-consommateurs.
La mise en place et la définition de ces concepts fondamentaux en écologie nous permet de comprendre que les écosystèmes possèdent une structure hiérarchisée sous différentes perspectives. Par exemple, si on s’intéresse à la densité d’une espèce (c’est-à-dire le nombre d’individu de cette espèce par unité de surface), on trouve toujours un plus grand nombre d’individus dans les niveaux trophiques les plus bas. Cependant, cette pyramide des nombres n’est pas vérifiée dans le cas des écosystèmes forestiers où on prend alors en compte la pyramide de biomasse (figure 1). Il existe également la pyramide d’énergie montrant la perte de l’énergie qui est transmise d’un niveau à l’autre. Ces pertes viennent de la production de déchets, de l’énergie utilisée lors de la récupération de nourriture, du métabolisme de l’organisme. L’énergie est produite au niveau inférieur et est disponible pour le niveau supérieur direct.
Les écosystèmes sont donc des réseaux dynamiques dépendant de l’hétérogénéité des forces d’interactions, d’énergie, de matière. On retrouve alors dans un écosystème des composantes piliers, qui ont un rôle essentiel, et des composantes secondaires ou annexes. En choisissant de classer les organismes par espèce, les écologues déterminent, dans des communautés, des espèces structurantes et des espèces interstitielles. Parmi les espèces structurantes on retrouve des espèces fondatrices ou des espèces clé-de-voûte.
Origine du concept d’espèce « clé-de-voûte »: l’expérience de R.T Paine
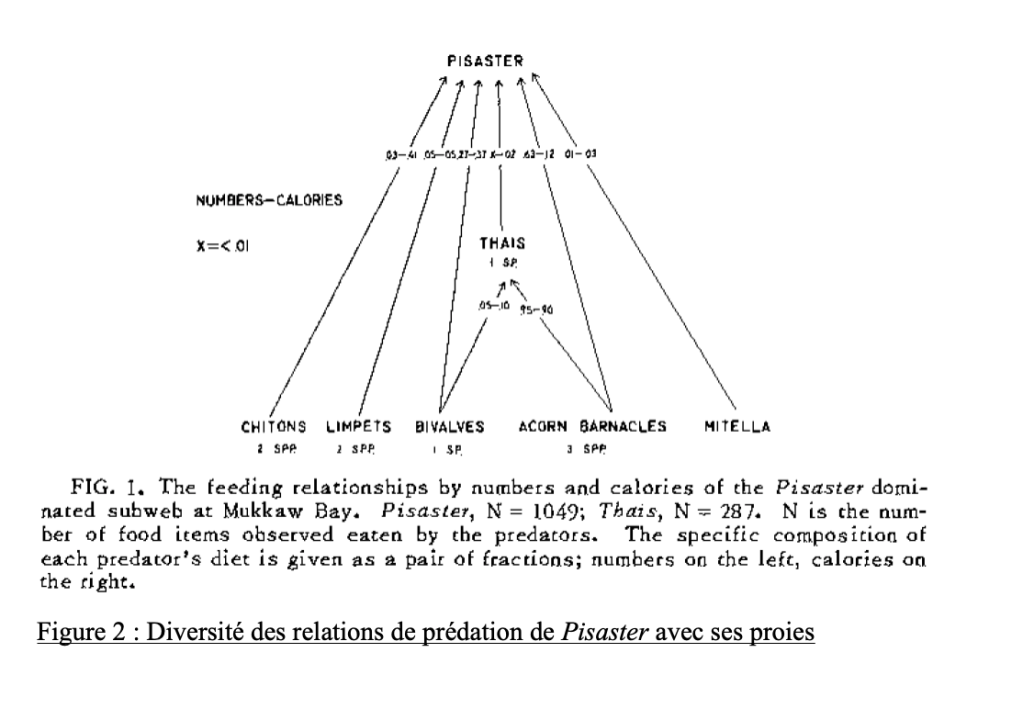
Dans les années 1960, Robert T. Paine étudie une communauté intertidale, sur une côte rocheuse, au sein de laquelle une étoile de mer (Pisaster sp.) est le prédateur de plusieurs mollusques. C’est en 1966 que Paine publie les résultats de son étude. L’expérience de Paine a pour but de déterminer quelle est l’importance de cette espèce d’étoile de mer concernant la composition et la stabilité des communautés de la zone intertidale.
Sur certaines zones, Paine retire toutes les étoiles de mer et empêche leur recolonisation. Sur d’autres, il laisse l’écosystème intact. Il compare ensuite l’évolution du peuplement des zones sans étoiles de mer et des zones laissées inchangées.
En l’absence de l’étoile de mer, il constate que les différentes espèces de mollusques entrent en compétition jusqu’à ce qu’il ne reste, à la fin, qu’une seule espèce de mollusque »dominante ». En revanche, la diversité des espèces de mollusques est maintenue en présence des étoiles de mer. On déduit de ces observations que l’étoile de mer Pisaster sp. doit avoir une prédation préférentielle pour les espèces dominantes dans les interactions de compétition entre mollusques, donc celles qui ont des populations plus abondantes – ce qui empêche alors l’exclusion compétitive des autres espèces de mollusques. Il parvient ensuite à établir du rôle structurant de la prédation de Pisaster sp. dans les communautés de la côte extérieure de l’état de Washington (figure 2).
R.T. Paine semble alors être le premier à utiliser le terme d’ espèce « clé-de-voûte » pour parler des espèces dont la présence est cruciale pour maintenir l’organisation et la diversité de la communauté écologique. Mais dans son emploi par Paine, le terme est en fait plus réducteur que la définition que nous lui attribuons. En effet, dans son article « A note on trophic complexity and community stability » paru en 1969 dans The American Naturalist Paine introduit la notion d’espèce « clef-de-voûte » :
« Within both these fairly or very complex systems the species composition and physical appearance were greatly modified by the activities of a single native species high in the food web. These individual populations are the keystone of the community’s structure, and the integrity of the community and its unaltered persistence through time, that is, stability, are determined by their activities and abundances »
La première définition donnée du concept d’espèce clef-de-voûte semble alors dépendre de la position de l’espèce concernée dans le réseau trophique. Une espèce clé-de-voûte est haut placée, il s’agit alors très probablement de prédateurs, et une espèce clé-de-voûte n’est alors sûrement pas un producteur primaire. C’est la position dans la chaine alimentaire ainsi que l’abondance relative et les activités qui établissent le rôle structurant et stabilisateur d’une espèce pour la communauté. Néanmoins, on note ici que Paine ne parle pas d’espèce « clé-de-voûte » mais de « populations individuelles ». Est-ce alors l’espèce, la population ou l’individu qui est « clé-de-voûte » ? N’y a-t-il d’espèces « clés-de-voûte » que chez les espèces prédatrices ?
Problèmes de la détermination de la composante « clé-de-voûte ».
On s’interroge désormais sur le niveau de lecture de la composante « clé-de-voûte » au sein d’un écosystème et de la relation entre la définition d’une espèce comme « clé-de-voûte » et sa position dans le réseau trophique. Dans le cas des étoiles de mer de Paine, l’espèce clé-de-voûte désigne clairement des prédateurs au sommet du réseau trophique qui, par leur présence et leur prédation préférentielle sur des espèces dominantes, favorisent la coexistence d’espèces potentiellement compétitrices. Le retrait des étoiles de mer entraine donc une chute de la biodiversité. Après l’expérience de Paine, d’autres écologues ont repris le terme de « clé-de-voûte » pour désigner des populations dont la présence détermine la structure d’une communauté bien que celle-ci ce soit pas un consommateur élevé dans la chaine alimentaire.
Dans le cas des loutres de Duggins, (1980) et Estes, Smith et Palmisano, (1978), l’absence de celles-ci entraine la prolifération des oursins qui, par leur broutage, modifient le paysage végétal sous-marin et donc la faune associée. L’action sur le peuple benthique passe par le surpâturage et les loutres ont un effet indirect sur la modification de l’environnement.
Avec les fourmis de feu Solenopsis germinata (Risch et Carrol, 1982) on se trouve dans le cas d’un prédateur non-spécialisé qui s’attaque à des herbivores faiblement compétitifs. Son absence n’entraine pas une régression de certaines populations ou la disparition d’espèce mais l’augmentation des effectifs et de la biodiversité au sein du peuplement d’arthropodes défavorables aux cultures.
Avec le concept de “ressources-clé” (Terborgh, 1986; Gilbert, 1980) puis celui d’« agenceurs écologiques » ( Jones, Lawton et Shachak, 1994), on assiste alors à une tentative de généralisation du concept de « clé-de-voûte » à des espèces non-prédatrices qui pourraient être à des piliers dans le maintien de la structure de leur communauté. On peut, par exemple, penser au castor qui est considéré comme un ingénieur écologique puisqu’en modulant directement ou indirectement la disponibilité des ressources ou l’architecture du milieu, il détermine la distribution des espèces d’un écosystème.
Alors, être une espère « clé-de-voûte » ne dépend pas exclusivement de la qualité intrinsèque de l’espèce (être herbivore, carnivore, macrophages …) ou de sa place dans le réseau trophique. Ces deux caractéristiques influencent l’importance de l’espèce dans la structure de la communauté mais ne sont pas des critères immuables quant à la détermination des espèces « clés-de-voûte ».
En effet, là où une espèce serait au centre de l’équilibre écosystémique dans un biotope précis, cette même espèce pourrait n’être qu’une espèce interstitielle dans un autre écosystème. Il s’agit alors de populations « clés-de-voûte », relatives à un fonctionnement et à une architecture d’une écosystème précis. Ce n’est pas parce que Pisaster sp. était « clé-de-voûte » dans le milieu étudié par Paine qu’elle le serait sur une autre côte. On remarque alors une particularité de la définition de l’espèce « clé-de-voûte » et une dépendance relative quant à l’écosystème étudié. Or on peut se demande s’il existe une distinction ontologique entre la population et l’espèce.
Au delà d’une distinction quantitative qui n’apporte rien au problème, pour des auteurs comme Dawkins, Hull ou Ghiselin, la différence entre population et espèce se trouve dans l’idée que la première est une partie de la seconde.
« Whether the relatively large units recognized by taxonomists as species evolve or whether much less extensive units such as populations are the effective units of evolution is an open question. In this paper when I use the term « species, » I intend to refer to those supraorganismic entities which evolve regardless of how extensive they might turn out to be »
De la différence ontologique entre espèce et population
Cependant, sans nier que population et espèce puissent être coextensives (dans le cas où il ne resterait qu’une population de l’espèce considérée par exemple), on peut soutenir l’idée qu’il existe bien une différence ontologique. On s’appuie alors sur la thèse de Mahner et Bunge (1997) dans leur ouvrage Foundations of biophilosophy. Ils tiennent une position conceptualiste reposant sur la distinction entre les organismes-individus réels et les classes-espèces abstraites.
« It holds that, in the systematic hierarchy, (a) only individual organisms are real, i.e., are concrete individuals and exist independently of the knowing subject (other material systems composed of organisms, such as biopopulations, communities, and social systems are real, too, but are disregarded here because they are not relevant to systematics); (b) species and the other taxa are concepts, though not arbitrary and useless ones, for they represent objective commonalities among organisms: they arenatural classes or, ideally, natural kinds. To paraphrase Christian Wolff: taxa nonexistunt, nisi in individuis (1740, §56). However, we shall see below that, in order to take evolution into account, the standard notion of a natural kind needs to be modified into the weaker notion of a biological kind. »
La relation entre ces deux taxons (espèce et population) est une relation d’inclusion ensembliste (⊂). La relation d’appartenance (∈) est seulement valide entre les organismes et la classe ultime (l’espèce ou « infima specie »). Les populations (« biopopulations ») sont composées d’organismes dans une relation de partie au tout. On distingue alors deux types de hiérarchies, une taxinomique qui regroupe tous les taxons (ordre, famille, genre, espèce, …) et une réelle composée d’entités matérielles (biome, écosystème, peuplement, métapopulation, population, individu/organisme). La classification taxinomique est phylogénétique, morphologique ou structurale et il y a donc une inclusion logique des classes les plus singulaires dans les taxons supérieures (par exemple, l’espèce est incluse dans le genre). La classification réelle est éco-évolutive, c’est-à-dire qu’elle est fonctionnelle et se base sur le niveau d’intégration des entités (la population est moins générale et est une partie de l’écosystème, qui lui est moins général et une partie d’un biome). La théorie évolutive tend à soutenir cet argument en plaçant l’espèce comme une unité taxinomique et la population comme une unité évolutive (d’où peut s’effectuer le phénomène de spéciation).
Problématique
Cette différence ontologique entre population et espèce éclaircit le concept de « clé-de-voûte » qui est un concept fonctionnel, se basant sur une réalité empirique et qui doit donc être rapporté à l’ontologie de la population plus qu’à celle de l’espèce telle que nous l’avons définie précédemment. Il s’agit alors pour l’écologue qui veut étudier le concept de « clé-de-voûte » d’adopter un regard plutôt systémique – c’est-à-dire considérant les ensembles organiques comme indécomposables en pièces, reliés les uns avec les autres et non pas comme un agrégat d’unités biologiques indépendantes – on privilégie alors la synécologie à l’autoécologie.
Se demander si une espèce clé-de-voûte est « clé-de-voûte » par essence revient désormais à se demander si une population peut être intrinsèquement définie comme « clé-de-voûte ». Les membres d’une population et cette population sont-ils nécessairement « clés-de-voûte » ? Nous nous demandons alors si l’attribut « clé-de-voûte » est essentiel ou accidentel.
La « clef de voûte », de la métaphore architecturale au rôle écologique et culturel de l’espèce.
Un emprunt au domaine technique de l’architecture
Le terme « clé-de-voûte » (keystone en anglais) est emprunté au vocabulaire de l’architecture et désigne initialement une pierre placée dans l’axe de symétrie d’un arc de cercle ou d’une voûte. Il permet de maintenir et bloquer les voussoirs de la structure. Ainsi, la clé-de-voute désigne un élément qui est à la fois central (essentiel) et qui assure un équilibre à un ensemble (ou un système). On peut alors comprendre par analogie le rôle essentiel des espèces clés-de-voute dans un écosystème. Il suffit d’enlever cette espèce pour constater l’effondrement de l’ensemble organisé de la biocénose. Cette notion semble alors bien définie et on ne voit aucun problème à cet emprunt de la « clé de voute » au vocabulaire technique de l’architecture par l’écologie. Cependant, on peut se demander si l’emprunt de cette métaphore résiste au passage du rouleau compresseur humain et de sa rôle impactant au sein des écosystèmes.
De la distinction de deux types de communautés
Il existe une diversité des situations d’interactions qui nous permet d’établir deux types de sociétés. La détermination de celles-ci dépend du niveau de perturbation de leur équilibre écosystémique par le facteur anthropique. Il y a des communautés vivantes non-humaines, vivant en harmonie avec leur milieu qu’on appellerait « sociétés naturalistes » dans le sens de la naturalité, comme sauvage, en opposition avec les communautés où la présence de l’Homme déséquilibre les interactions biotiques. Cette dichotomie soulève une question que l’on peut formuler ainsi : la notion de « clé-de-voûte » ne s’applique-t-elle qu’à des espèces sauvages – donc aux sociétés « naturalistes » – ou peut-elle être une notion utilisée dans nos sociétés humanisées comprenant un grand nombre d’espèces domestiquées ? Les écosystèmes considérés lorsqu’on parle d’espèce « clé-de-voûte » sont-ils seulement sauvages ou peut-on aussi appliquer ce concept à des sociétés humaines ?
Néanmoins, nous pouvons nous demander comment légitimer cette séparation entre une société « de la nature » et une société « de la culture » que nous venons d’établir ? Comment différencier espèces sauvages et espèces domestiques ? Cette distinction entre sauvage et domestique n’est pas sans rappeler « l’opposition faite par les géographes entre écoumène et érème, entre les lieux que les hommes fréquentent au quotidien et ceux où ils s’aventurent plus rarement » et reste très discutée par les philosophes de par la divergences de compréhension de ce couple terminologique au sein des différentes cultures, comme le montre Philippe Descola.
La création de cette dichotomie entre espaces sauvages et espaces humanisés exerce un pouvoir de réduction sur le champ d’application du concept de « clé-de-voûte », qui semble inapplicable aux sociétés anthropiques dont les relations biotiques et abiotiques semblent avoir été bouleversées par la colonisation du milieu par les humains. Les écosystèmes divers et variés qui ont permis l’émergence et le maintien d’une grande diversité de systèmes sociaux humains et non-humains voient leur endurance altérée par les changements sans précédent de l’Anthropocène. Dans les sociétés civilisées et urbanisées, peut-on encore trouver des espèces « clé-de-voûte » ?
Émergence d’une nouvelle conception de l’espèce « clé-de-voûte »
Lors du colloque de Villejuif « Le symbolisme des animaux » (2003) a été discuté le concept de « l’animal de civilisation ».
« Un animal (ou groupe d’animaux) serait dès lors une “espèce clef de voûte culturelle” lorsque la société concernée a élaboré à son propos un système de récits, croyances et pratiques dont l’étude permet d’approcher l’identité culturelle de la société à travers sa relation à son environnement naturel. »
La dichotomie entre communautés anthropiques et communautés naturalistes propose une nouvelle généralisation du concept de l’espèce « clé-de-voûte » encore plus marquante que celles proposées par les écologues du vingtième siècle, qui reposaient sur des différences d’échelles ou de rôle trophique. Désormais, on pose l’existence d’une « clé-de-voûte culturelle » qui va au-delà de la population d’un écosystème et ne s’applique pas uniquement à des organismes vivants réels mais aussi à des créatures extraordinaires, des mythes – l’espèce prend désormais des aspects divers allant de l’animal sauvage au monstre en passant par l’animal domestique, la curiosité hybride, la déité ou le symbole.
L’espèce « clé-de-voûte » , du naturel au culturel.
L’espèce clé-de-voûte dans les sociétés sauvages
Nous avons vu en introduction qu’une espèce clé-de-voûte, en écologie, était une espèce ayant un effet très important, voire disproportionné sur son environnement en vue de ses effectifs ou de sa biomasse. C’est une espèce dont la présence est indispensable à l’équilibre de l’écosystème. Son activité et son abondance déterminent l’intégrité de la communauté, sa persistance, sa stabilité. La disparition de cette espèce « clé-de-voûte » entraine des changements fonctionnels majeurs.
Historiquement, l’importance des espèces a d’abord été évaluée par leur place dans le réseau trophique. Le concept de « clé-de-voûte » redistribue les cartes : un herbivore, une plante, un champignon peuvent eux aussi être des espèces « clés-de-voûte ». D’ailleurs, le rôle clé-de-voûte d’une espèce peut se présenter de différentes façons et on peut classer les espèces « clés-de-voûte » en différents groupes non-exclusifs. Il y a des organismes ayant un rôle dominant au sein de l’écosystème, des espèces fournissant les ressources indispensables à un nombre conséquent d’organismes, des espèces co-dépendantes dont la disparition d’un des mutualistes entrainerait la disparition de l’autre, et enfin, les ingénieurs écologiques qui sont des organismes modifiant physiquement l’écosystème et fournissant par exemple un habitat à d’autres espèces, habitat qui ne serait pas disponible autrement.
L’attribut « clé-de-voûte » est donc assigné à une population en fonction de son rôle dans l’écosystème. Or cette population si elle est ainsi dans l’écosystème, c’est de façon « naturelle ». Ce n’est ni un choix, ni un but que les organismes se seraient fixé d’atteindre à tout pris. On peut alors concevoir la définition fonctionnelle de l’espèce « clé-de-voûte » comme étant un attribut essentiel d’une population au sein d’un écosystème. Si cette population venait à perdre son rôle de « clé-de-voûte » c’est que l’écosystème aurait changé et donc que la population aussi que ce soit dans sa nature, dans son effectif, sa biomasse. La population clé-de-voûte est essentiellement centrale et le moyen de lui faire perdre de cette importance serait de l’éradiquer. Or alors, elle ne serait plus. Voilà pourquoi on peut soutenir que l’espèce « clé-de-voûte » l’est par essence.
L’espèce clé-de-voûte dans les sociétés humaines
De plus, dans les sociétés anthropiques, l’espèce clé-de-voûte culturelle peut aussi prendre plusieurs formes. On peut tout d’abord qualifier d’espèce « clé-de-voûte » des espèces domestiquées qui ont un rôle central et important dans le fonctionnement de la société humaine. Ces espèces « clé-de-voûte » peuvent donc être déterminés par leur rôle structurant d’un point de vue social, culturel ou même vital (dans l’alimentation par exemple).
Éric Garine dans ses Notes de terrain sur une civilisation agraire du Nord du Cameroun (Duupa, Massif de Poli) a étudié le symbolisme et l’importance du bétail dans des sociétés d’agriculteurs des montagnes du nord du Cameroun. Il y affirme alors que « toutes les espèces connues dans une société particulière n’y jouent pas des rôles équivalents dans la définition des principes idéologiques sur lesquels repose sa structure sociale, les fondements de son identité culturelle, ou son fonctionnement écologique. Certaines espèces sont indispensables à la définition même de la société et à sa reproduction. » comme c’est le cas pour le cochon pour la société Tsembaga de Nouvelle-Guinée ou le bétail dans les sociétés pastorales d’Afrique de l’Est dont la vie sociale est indissociable de l’élevage. Il note également qu’il semblerait que pour le peuple Nuer, le zébu est un rôle clef et que sa disparition entrainerait un effondrement de la société, aussi bien d’un point de vue matériel puisqu’ils ne pourraient plus assurer leur subsistance que culturel puisque le zébu se retrouve aussi dans leurs mythes, il a également le rôle de monnaie (notamment lors des unions et mariages). Ainsi, on constate une relation vitale entre la société humaine et l’espèce animale en question – au-delà du rôle écologique et matériel qui est pourtant primordial du point de vue de la subsistance, c’est le rôle social de l’espèce clef de voute culturelle qui importe puisqu’elle se place comme source de culture et comme un média dans les relations sociales. Elle est ainsi centrale et permet l’équilibre des sociétés concernées.
Quant à l’espèce « clé-de-voûte culturelle » non-vivante, comme une déité par exemple, c’est une espèce « clé-de-voûte » par essence dans la mesure où elle a été formée pour répondre au besoin d’équilibre, pour combler une place centrale manquante dans cette société. La création de l’espèce est en elle-même accidentelle car désirée par les agents de la communauté humaine, mais l’espèce est essentiellement « clé-de-voûte » sinon elle ne serait pas. Cependant, on s’éloigne du concept tel qu’il a été formulé par R.T. Paine – même si on raisonne analogiquement – puisque qu’une divinité est immuable, et il serait alors impossible de voir son réel impact sur la communauté en la faisant disparaitre. Il semble alors que cette espèce « clé-de-voûte culturelle » ne puisse émerger qu’en quasi-simultanéité avec la communauté qui la crée.
Il devient alors essentiel à l’anthropologue, s’il veut saisir l’ontologie de l’être humain, de s’intéresser aux espèces non-humaines, notamment aux espèces clés-de-voûte culturelles dont nous avons vu des exemples précédemment. En effet, l’anthropologie comme l’écologie ou l’éthologie semblent bien plus capables de décrire que de définir les espèces qu’elles étudient. Alors en s’intéressant à ce que l’Homme produit et ce qui unit les Hommes entre eux, en structurant leur vie en société, l’anthropologue s’approche un peu plus du noyau de la nature humaine qu’il cherche à trouver. C’est l’existence de ces espèces clés-de-voûte culturelles dans des sociétés animistes et totémistes qui poussent aujourd’hui les anthropologues à introduire l’ontologie dans l’anthropologie.
Une lecture naturaliste des sociétés humaines : l’hyperkeystone humaine
En sortant de la dichotomie précédente et en ne différenciant plus les communautés en fonction de la présence ou de l’absence de l’humain – c’est-à-dire en étudiant les sociétés humaines avec le regard d’un écologue étudiant des sociétés naturalistes, on constate que l’Homo sapiens a une place centrale, éminente et influente sur son environnement.
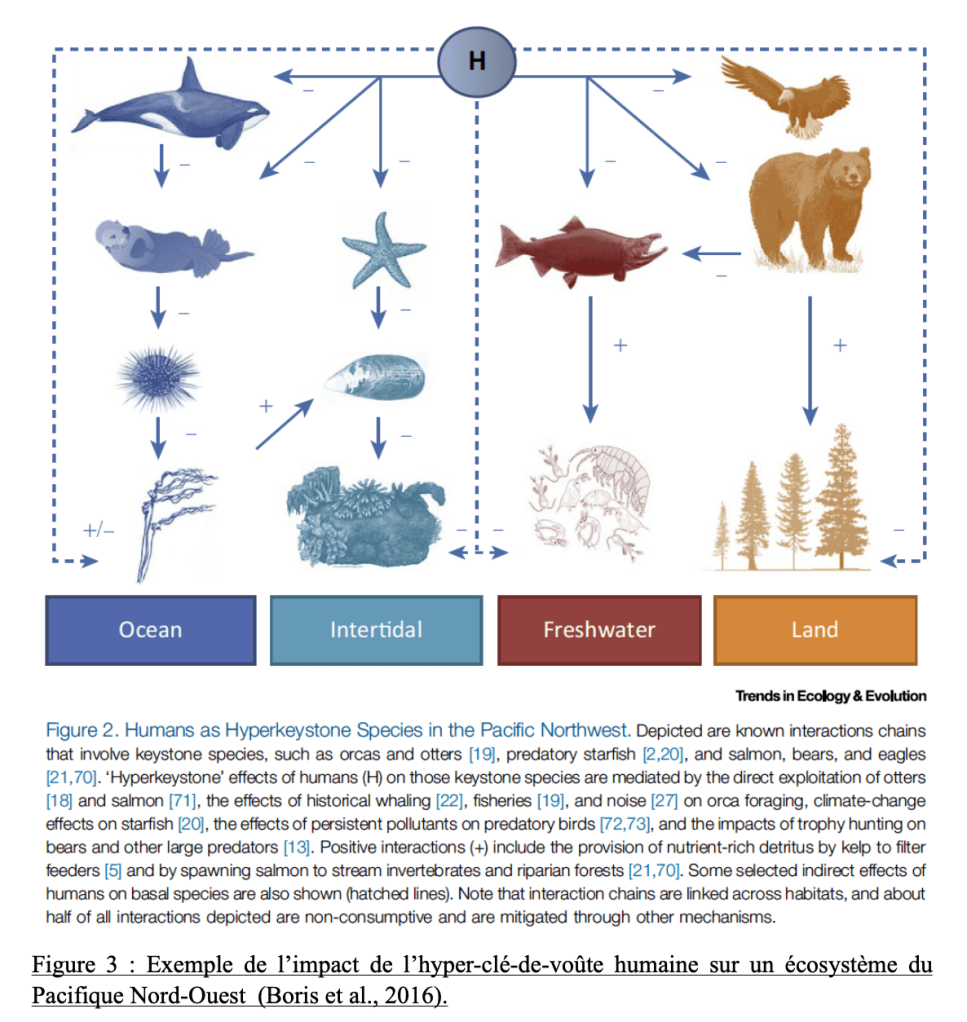
L’Homo sapiens comme hyperkeystone, en français littéralement « hyper-clé-de-voûte », c’est l’idée que l’espèce humaine « dirige des chaînes d’interaction complexes en affectant d’autres acteurs clé-de-voûte au sein des différents habitats. » L’homme se fait « clé-de-voûte » au-dessus des « clés-de-voûte » et réorganise les réseaux trophiques en modifiant la position des clefs de voûte traditionnelles. Ce rôle se rapproche tout autant de celui de « hyperparasite », une espèce qui affecte plusieurs autres espèces clés au sein d’habitats différents (Boris et al., 2016). L’Homme n’est plus une espèce « clé-de-voûte » traditionnelle puisque ce n’est pas à l’échelle d’une population ou d’un écosystème qu’il se rend indispensable mais bien dans tous les écosystèmes qu’il occupe. Ce n’est plus un rôle « naturel » mais il se rend nécessaire et s’impose à toutes les composantes des biomes envahis dans le but de contrôler, maitriser son environnement et l’exploiter, aux dépens du reste des composantes de l’environnement (exemple : figure 3)
Cette notion de « hyper-clé-de-voûte » interroge sur une éventuelle échelle ontologique ? Y a-t-il des espèces qui sont plus « clés-de-voûte » que d’autres ? L’Homme est-il plus « clé-de-voûte » que les populations « clés-de-voûte » qu’il surplombe ? De plus, l’appropriation par l’Homo sapiens du rôle de « clé-de-voûte » entraine-t-il la perte de ce rôle par les espèces « clé-de-voûte » traditionnelles ?
La question est d’ordre ontologique, car si l’Homme parvient à s’imposer comme hyperkeystone dans un écosystème sans en changer profondément la nature, les effectifs, la biomasse, le paysage – par exemple s’il s’installe dans cet écosystème comme une « hyper-clé-de-voûte » potentielle mais qu’il n’actualise pas ce potentiel en modifiant les rapports existants – alors l’attribut « clé-de-voûte » retiré aux espèces devient un attribut accidentel et non pas essentiel. Il se pourrait que toutes les espèces soient alors toujours « clé-de-voûte potentielle » et que seul un certain contexte permette l’actualisation de ce potentiel.
Ainsi, en vocabulaire aristotélicien, les espèces admettent en puissance l’attribut « clé-de-voûte » qui ne peut-être actualisé qu’en fonction du lieu (écosystème) dans lequel elles se trouvent. Cet attribut appartient donc à la forme de ce type de substance, l’espèce et c’est donc un attribut essentiel. Les espèces clé-de-voûte le sont par essence. Or pour Aristote, si c’est une disposition naturelle, ce qui est le cas pour toutes les puissances qui appartiennent à la forme des vivants, alors il est nécessaire pour cette espèce de se trouver naturellement dans tel écosystème qui permet l’actualisation de son potentiel rôle « clé-de-voûte ». Cela signifierait qu’à chaque espèce on devrait associer un écosystème donné. Pour Aristote, il faut se diriger vers l’excellence à partir de toute disposition naturelle. Si l’on prend le concept de « clé-de-voûte » au sens faible comme signifiant « être nécessaire à tel ou tel écosystème », le fixisme aristotélicien ne permettant pas de lire une forme d’évolution au sein des populations. Aristote n’avait découvert que quatre cent quatre-vingt quinze espèces animales, et il était loin du paradigme moderne de l’adaptation des populations aux milieu par les mécanismes évolutifs (mutation, dérive génétique, sélection naturelle, flux de gènes…).
Il ne faut oublier que la base de la définition du concept de « clé-de-voûte » est une base empirique et relativement moderne, qui peut trouver soutien et appui dans la théorie darwinienne, la génétique moderne, l’écologie fonctionnelle et la biologie des populations. Conclusion
Conséquences éthiques et pratiques
Du symbolisme engendré par ce concept
Qu’il soit « clé-de-voûte » culturelle ou écologique, l’organisme clé-de-voûte s’impose dans nos esprits comme un symbole, animiste, religieux, spirituel ou environnemental. Par exemple, le peuple des Punan de Bornéo utilise la réponse adaptative des sangliers (Sus barbatus barbatus) à la phénologie des Dipterocarpaceae, famille d’arbre dominante dans leurs forêts. Cette espèce de sanglier est d’abord une espèce clé-de-voûte écologique en ce qu’elle permet de déterminer l’état des dégradation des forêts de Dipterocarpaceae. En effet, les sangliers ont développé une réponse adaptative à la fructification massive des Dipterocarpaceae, ce sont des omnivores étendus ce qui leur permet de trouver des ressources alternatives lorsqu’il y a moins de fruits de Dipterocarpaceae; les femelles peuvent avoir deux portées s’il y a abondance de nourriture disponible; les pics de naissance sont synchronisés avec les périodes de chute des fruits etc. L’écologie de ces sangliers migrateurs témoignent donc de leur niveau d’adaptation élevé. Par de nombreuses facettes (aération des sols par le fouissage, dispersion des graines lors des migrations etc), le sanglier contribue à l’entretien et à la dynamique des forêts.
Néanmoins, c’est aussi une espèce « clé-de-voûte culturelle » puisqu’il joue un rôle central dans la vie rituelle et la cosmologie de la société. La viande de sanglier est un aliment qui possède une grande valeur culturelle dans les sociétés de chasseurs-collecteurs, il s’agit d’un met de choix. De plus pour les Punan, le sacrifice des sangliers constitue un acte charpentant les périodes de guerres. Il s’agit d’un animal sacré dont on consomme la viande avec respect et parcimonie. C’est également un animal admiré pour sa force, son faible nombre de prédateurs natures et pour sa capacité à collaborer avec d’autres espèces. Pour les Punan, le sanglier possède une propriété surnaturelle, mythe largement influencée par son comportement migratoire : il a le rôle de messager. On remarque alors que ce sont les propriétés écologiques du statut de « clé-de-voûte » du sanglier qui lui procure cette importance culturelle. Le concept de « clé-de-voûte », en pouvant se transposer » du naturel au culturel », permet aux espèces concernées de passer d’un niveau d’être matérielle et empirique à un niveau d’être symbolique, idéel. Ce concept permet une transition ontologique.
De la théorie à la pratique
En attribuant la dénomination de « clé-de-voûte » (culturelle ou écologique) à une espèce, on tend à faire s’élever cette espèce dans une sorte d’échelle ontologique au risque de ne voir nos regards se concentrer sur elle et oublier le reste de l’écosystème sans lequel elle ne pourrait subsister. Cependant nous avons vu qu’écologiquement, il s’agit pour nous de parler de populations « clés-de-voûte » et que cette définition n’est pas la plus objective, de par les différentes utilisations de ce même terme pour désigner plusieurs formes d’espèces structurantes. Une fois cette théorie de l’espèce clé-de-voûte mise en place et nous encadrant, il faut désormais s’intéresser à son application dans les champs pratiques de l’écologie : l’ingénierie et la restauration écologique et la biologie de la conservation.
La biologie de la conservation des espèces est présentée par Robert Barbault comme une discipline de gestion de crise qui doit identifier les populations en déclin et les espèces en danger pour déterminer les causes de leur déclin puis qui doit proposer des moyens d’y remédier. Ainsi c’est à partir de notre connaissance des structures des écosystèmes qu’on peut prévoir et comprendre les risques « d’extinction en cascades ». La restauration est une partie de la biologie de la conservation puisqu’il s’agit alors de la mise en oeuvre sur le terrain des moyens trouvés par les chercheurs en éco-conservation. Connaitre la structure des écosystème permet alors d’augmenter l’efficacité des plans d’actions en ciblant les projets de restaurations sur les espèces clé-de-voûte qui sont centrales. On pourra également déduire de cette connaissance les contraintes à respecter, comme le fait de restaurer les populations d’espèces qui conditionnent la présence de celles que l’on souhaitent voir s’épanouir à nouveau. Le meilleur moyen serait donc de se focaliser sur les espèces « clés-de-voûte » écologiques ou d’affiner « l’approche fonctionnelle en subdivisant les niveaux trophiques en sous-ensembles plus précis, les guildes ou groupes fonctionnels. ». Cependant « (…) les deux conceptualisations impliquent différents modèles de structure de la communauté et nécessitent donc différentes stratégies de conservation. Si beaucoup ou la plupart des espèces sont d’une importance similaire, tout effort pour sauver seulement quelques clefs de voûte échouera inévitablement à protéger le reste. ». Le rôle du concept de « clé-de-voûte » écologique doit donc être manié avec un certain esprit critique en biologie de la conservation et seul l’établissement d’une définition universelle et partagée permettra l’élaboration de lois concernant la sauvegarde et la conservation des espèces. Or l’Homme, en tant qu’acteur principal de la conservation, reste biaisé par la multitude de définitions et d’utilisations du concept de « clé-de-voûte » et tout le symbolisme en découlant. Il ne peut pas faire un choix arbitraire quant à la répartition des ressources budgétaires en écologie de la conservation. L’Homme influencé par son héritage culturel et perdu dans un ensemble de définitions vagues et imprécises n’a pas encore en mains toutes les clefs pour sauvegarder au mieux les écosystèmes en péril.
Le rôle de l’Homme
Mark Dion dans l’une de ses expositions, a présenté son oeuvre Survival of the Cutest (2004) (figure 4). Il s’agit d’une brouette sur laquelle il a écrit les faits les plus importants concernant l’extinction des espèces et dans laquelle il a placé des peluches d’animaux pour enfants. Son message peut être le suivant : sommes-nous en train de privilégier la sauvegarde de certaines espèces au profit des autres en fonction d’un critère purement esthétique ?
Au-delà de l’anthropocentrisme, on peut évoquer un certain ethnocentrisme autour de la conservation des espèces-symboles des nations, ou des peuples. Les américains seraient bien plus effondrés par la disparition de leur pygargue (aigle à tête blanche américaine) que nous, français lointains nous le serions – et inversement, les américaines ne semblent pas porter grand intérêt à la disparition de l’ours des Pyrénées ou des cigognes d’Alsace. Parce que c’est l’Homme qui est l’agent conservateur, les espèces « clé-de-voûte culturelles » semblent bénéficier d’un traitement de faveur dans la sauvegarde de espèces (en Chine on peut être condamné à mort si l’on tue un panda).
Ainsi, c’est encore une démonstration du rôle de l’Homo sapiens comme « hyperkeystone ». Il se place en maitre et partie de l’écosystème auquel il s’intéresse. Chacun de ses choix de vie finit par avoir des impacts profonds sur les espèces clés des différents habitat. De plus, n’oublions pas que ce que nous ne présentons ici n’est qu’une facette des répercussions de l’ère Anthropocène sur la biosphère.
Au regard d’une définition traditionnelle du concept « clé-de-voûte », les espèces – ou populations – auxquelles nous attribuons cette qualité semblent alors se poser comme essentiellement « clé-de-voûte ». Cependant, l’évolution du concept dans des perspectives modernes et culturelles, l’apport des découvertes scientifiques et surtout l’omnipotence de notre espèce Homo sapiens sur la quasi-totalité de la biosphère et des écosystèmes tend à en faire un caractère muable et contingent – dont nous serions au final à la fois le créateur et le produit – l’humain comme maitre et seule véritable espèce clé-de-voûte .
Bibliographie
BARBAULT Robert (1997)
Biodiversité. Introduction à la biologie de la conservation, Hachette
BARBAULT Robert (1995)
« Le concept d’espèce-clé de voute en écologie de la restauration : clé… ou impasse ? », Nature – Sciences – Sociétés Hors-série
ELTON Charles (1927)
Animal Ecology, New York, Macmillan Co.
WORM Boris et PAINE Robert T. (2016)
« Humans as a Hyperkeystone Species », Trends in Ecology & Evolution , Volume 31
DELORD Julien (2010)
L’extinction d’espèce, histoire d’un concept & enjeux éthiques Paris : Muséum national d’Histoire naturelle, 2010, 691 p. (Archives ; 14).
DESCOLA Philippe (2005)
Par-delà nature et culture, Paris, Gallimard, coll. Folio essais
DESCOLA Philippe (2004)
« Le sauvage et le domestique », Communications, 76, 2004. Nouvelles figures du sauvage.
DOUNIAS Edmond, MOTTE- FLORAC Élisabethet DUNHAM Margaret (2007)
Le symbolisme des animaux : L’animal, clef de voûte de la relation entre l’homme et la nature ?, IRD Éditions, Collection Colloques et Séminaires, Paris
HULL David L. (1978)
« A matter of individuality », Philosophy of Science n°45 pp. 335-360
MAHNER Martin BUNGE Mario (1997)
Foundations of biophilosophy., Springer
MILLS, SOULÉ, DOAK (1993)
« The Keystone-Species Concept in Ecology and Conservation », BioScience, Vol. 43, No. 4
PAINE Robert T. (1966)
« Food Web Complexity and Species Diversity », The American Naturalist, Vol. 100, No. 910
PAINE Robert T. (1969)
« A Note on Trophic Complexity and Community Stability », The American Naturalist, Vol. 103, n° 929
TIRARD Claire, ABBADIE Luc, LALOI David et KOUBBI Philippe(2016)